
尨巕椡僔僗僥儉尋媶奐敪帠嬈媦傃尨巕椡婎慴婎斦愴棯尋媶僀僯僔傾僥傿僽丂惉壥曬崘夛帒椏廤
僋儕僾僩價僆僔僗偲儕儞僋偟偨曻幩慄懴惈婡峔偺夝柧尋媶
乮尋媶戙昞幰乯墱揷棽丂崺拵壢妛尋媶椞堟
乮嵞埾戸愭乯崙棫戝妛朄恖幁帣搰戝妛
乮尋媶奐敪婜娫乯暯惉俀侽擭搙乣俀俀擭搙
侾丏尋媶奐敪偺攚宨偲偹傜偄
丂尨巕椡敪揹傪廳梫側僄僱儖僊乕嫙媼尮偲偟偰偄傞変偑崙偱偼丄曻幩慄偺惗懱塭嬁偼崙柉偺埨慡丒埨怱傪妋曐偡傞偨傔偺嵟廳梫壽戣偱偁傞丅傑偨丄巰場偺戞堦埵偑乬偑傫乭偱偁傞傢偑崙偱偼丄PET丄PETCT摍偺曻幩慄傪梡偄偨崅搙堛椕恌抐偍傛傃廳僀僆儞摍偺曻幩慄傪梡偄偨偑傫帯椕偑恑揥偟偰偍傝丄曻幩慄惗懱塭嬁偺昡壙偲曻幩慄杊岇偼幮夛揑偵傕廳梫偱偁傝嬞媫傪梫偡傞壽戣偱偁傞丅廬棃偺曻幩慄惗懱塭嬁尋媶偼丄曻幩慄崅姶庴惈偵娭梌偡傞婡彉夝柧偵懳偡傞尋媶偑拞怱偱偁傝丄暘巕惗暔妛揑庤朄傪梡偄偰條乆側曻幩慄墳摎場巕偑柧傜偐偵偝傟偮偮偁傞丅偟偐偟丄曻幩慄崅懴惈場巕偍傛傃婡彉偺慡梕偑枹偩晄柧側揰偑懡偄丅
丂惗暔偺曻幩慄崅懴惈偺暘巕婡峔偵偮偄偰偼丄曻幩慄懴惈嵶嬠D. radiodurans傪拞怱偲偟偰丄摿偵DNA偺廋暅婡峔偵偮偄偰偺徻嵶側夝愅偑恑傔傜傟偰偒偨丅D. radiodurans偺崅偄曻幩慄懴惈偼姡憞偵揔墳偟偰敪払偟偨傕偺偲峫偊傜傟偰偍傝丄姡憞懴惈偲曻幩慄懴惈偺婡峔偵偼嫟捠揰偑懡偔擣傔傜傟傞丅懡嵶朎惗暔偵傕嬃偔傋偒姡憞懴惈傪帩偮傕偺偑偁傝丄惗懱撪偺悈暘傪傎傏姰慡偵幐偭偰傕惗柦傪堐帩偟丄嵞傃媧悈偡傟偽妶摦傪嵞奐偱偒傞丅偙偺惗柦尰徾傪僋儕僾僩價僆僔僗偲尵偄丄偦偺擻椡傪帩偮惗暔乮僸儖僈僞儚儉僔丄僋儅儉僔丄僽儔僀儞僔儏儕儞僾摍乯偼曻幩慄偵懳偟偰傕崅偄懴惈傪帩偮偙偲偑抦傜傟偰偄傞丅椺偊偽丄僱儉儕儐僗儕僇偺応崌丄姡憞梒拵偵兞慄傪7,000Gy徠幩屻丄悈偵栠偟偰梒拵偼慼惗偟乮Watanabe et al.,2006a乯丄廳棻巕慄乮僱僆儞乯傪200 Gy 徠幩偟偨応崌偱傕梒拵偼慼惗偟鍖偵曄懺偟偨乮Watanabe et al.,2006b乯丅幚嵺丄僱儉儕儐僗儕僇梒拵偺姡憞乮扙悈乯偵敽偭偰曻幩慄懴惈偑彊乆偵忋徃偟偰偄偔乮Nakahara et al., 2008乯丅偟偐偟丄偦偺暘巕婡峔偵偮偄偰偼傎偲傫偳暘偐偭偰偄側偄丅僋儕僾僩價僆僔僗摦暔偺拞偱嵟傕戝宆偺僱儉儕儐僗儕僇偼丄嵟嬤丄尋媶戙昞幰偵傛偭偰儌僨儖惗暔壔偝傟丄偦偺堚揱巕忣曬傕拁愊偟偮偮偁傞丅偮傑傝丄僱儉儕儐僗儕僇偺僋儕僾僩價僆僔僗偺暘巕婡峔傪夝柧偡傞偙偲偼丄曻幩慄懴惈偺婡彉傪柧傜偐偵偡傞偙偲偵懠側傜偢丄曻幩慄旐偽偔偵懳偡傞帠慜杊岇嶔偺僸儞僩偵宷偑傞偱偁傠偆丅
俀丏尋媶奐敪惉壥
丂僋儕僾僩價僆僔僗忬懺偺僱儉儕儐僗儕僇偼曻幩慄偵懳偟偰崅偄懴惈傪帵偡丅偙偺暘巕婡彉傪夝柧偡傞偨傔丄姡憞媦傃曻幩慄僗僩儗僗偵傛偭偰桿摫偝傟傞堚揱巕傪栐梾揑偵挷嵏偟偨丅傑偢姡憞夁掱偺俁抜奒乮0h, 12h偲36h乯偱僱儉儕儐僗儕僇梒拵偐傜cDNA偑摼丄EST僨乕僞儀乕僗傪峔抸偟偨丅偦偺姡憞懴惈娭楢EST僨乕僞儀乕僗傪夝愅偟偨偲偙傠丄慡晹偱15,054僋儘乕儞偑4,807堚揱巕僋儔僗僞乕偵暘椶偝傟偨丅偦偺敿暘偼怴婯堚揱巕偱偁傞偲峫偊傜傟傞丅儐僗儕僇偺摿挜偲偟偰丄僿儌僌儘價儞偑懡偔丄慡堚揱巕偺7.5%傪愯傔偰偄偨丅僋儘乕儞悢偵婎偯偄偨堚揱巕偺梊憐敪尰僷僞乕儞傪挷傋丄姡憞偵傛偭偰嫮偔桿摫偝傟丄姡憞懴惈偵娭傢偭偰偄傞偲巚傢傟傞堚揱巕偵拲栚偟偨丅摿偵僔儍儁儘儞婡擻側偳傪帩偮LEA僞儞僷僋幙丄HSPs丄偦偟偰巁壔僗僩儗僗娭楢堚揱巕偺僌儖乕僾偼姡憞偲嫮偔憡娭傪帵偟偨丅嵶朎偑姡憞偡傞帪偵嵟傕嫼埿偲側傞偺偑妶惈巁慺偺敪惗偵傛傞惗懱暘巕偺巁壔偱偁傞丅僨乕僞儀乕僗夝愅偵傛偭偰丄僱儉儕儐僗儕僇梒拵偼姡憞偺夁掱偱懡偔偺峈巁壔場巕傪崌惉偟丄妶惈巁慺偺嫼埿偵懳峈偟偰偄傞偙偲偑暘偐偭偰偒偨丅偟偐偟丄梊杊嶔傪島偠偰偄傞偵傕偐偐傢傜偢丄僋儕僾僩價僆僔僗忬懺偺僱儉儕儐僗儕僇梒拵偺DNA偼愗抐偝傟偰偍傝丄慼惗屻偵廋暅偡傞昁梫偑偁傞偙偲偑暘偐偭偨丅偙偺EST僨乕僞儀乕僗偺夝愅偵娭偡傞惉壥偼JBC (IF:6.4) 偵搳峞丄庴棟偝傟偨乮Cornette et al., 2010乯丅僱儉儕儐僗儕僇偺崅偄曻幩慄懴惈偼丄乽惗懱惉暘偺懝彎傪夞旔偡傞婡峔乿偲乽懝彎傪庴偗偨偲偒偺廋暅婡峔乿偵傛傞傕偺偲峫偊傜傟丄偦傟傜偼姡憞僗僩儗僗偵揔墳偟偰敪払偟偨傕偺偲峫偊傜傟偨丅偦偙偱丄僋儕僾僩價僆僔僗夁掱偱偺DNA懝彎偵偮偄偰僐儊僢僩傾僢僙僀朄傪梡偄偰夝愅偟偨偲偙傠丄幚嵺崅偄昿搙偱DNA嵔偺愗抐偑妋擣偝傟偨偑丄嵞悈榓屻偵偦傟傜偺懝彎偼廋暅偝傟偨乮恾侾乯丅
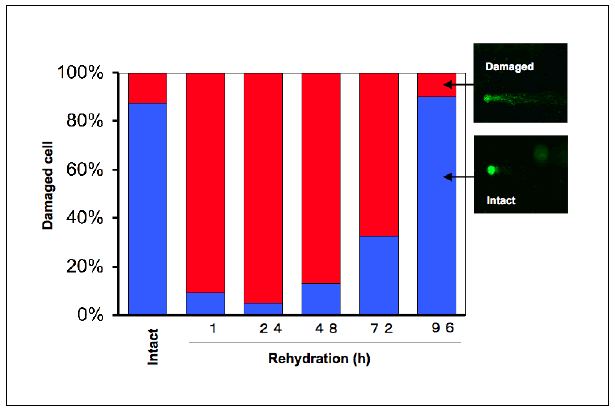
恾侾丂僱儉儕儐僗儕僇偺僋儕僾僩價僆僔僗乮姡憞僗僩儗僗乯偵敽偆DNA懝彎偲廋暅
掕朄偱姡憞偝偣偨梒拵傪悈偵栠偟丄堦掕帪娫屻偵夝朥偟偰帀朾懱嵶朎傪揈弌偟偨丅嵶朎傪屌掕偟丄傾儖僇儕忦審偺僐儊僢僩傾僢僙僀偵傛傝DNA懝彎傪掕検偟偨丅
丂師偵丄姡憞僗僩儗僗偲曻幩慄僗僩儗僗偵傛傞DNA懝彎婯柾偺夝愅傪峴偭偨丅妶摦忬懺偺僱儉儕儐僗儕僇梒拵偵50 MeV 4He (LET亣=16.2 keV/㎛)丂僀僆儞傪徠幩偟偨丅慄検偼敿悢塇壔慾奞慄検(70 Gy)偍傛傃敿悢鍖壔慾奞慄検(160 Gy)偲偟偨丅僱儉儕儐僗儕僇梒拵偵4He傪徠幩偟丄12帪娫屻偵僐儊僢僩傾僢僙僀傪峴偭偨偲偙傠丄慄検埶懚揑偵DNA偑懝彎偟偰偄偨 (恾俀)丅堦曽丄姡憞偐傜慼惗偟偨梒拵偺DNA傕懝彎偟偰偍傝丄偦偺掱搙偼70 Gy偺4He傪徠幩偟偨応崌偲嬤帡偟偰偄偨丅偄偢傟偺応崌傕丄傎偲傫偳偺嵶朎偑DNA偵懝彎傪晧偭偰偄偨偵傕偐偐傢傜偢丄傾億僩乕僔僗傗僱僋儘乕僔僗偵傛傞嵶朎巰偼傢偢偐偟偐娤嶡偝傟側偐偭偨丅曻幩慄徠幩嬫偍傛傃姡憞張棟嬫偲傕偵丄48帪娫栚傑偱偼懝彎DNA偺妱崌偵曄壔偼側偐偭偨偑丄96帪娫栚偵偼尭彮偟偨丅
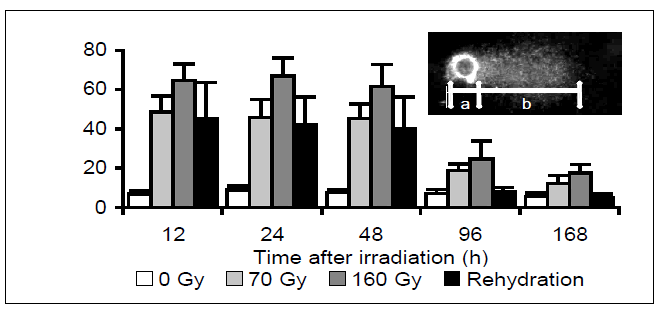
恾俀丂僱儉儕儐僗儕僇偺曻幩慄徠幩屻(0-160 Gy)偍傛傃姡憞偐傜慼惗屻(Rehydration)偺懝彎DNA偺検揑曄壔丅懝彎DNA(%)= b/(a+b)x100
丂妶摦忬懺偺僱儉儕儐僗儕僇梒拵偵丄敿悢塇壔慾奞慄検(70 Gy)偺Co-60僈儞儅慄偲摨慄検偺4He僀僆儞傪徠幩偟偨丅徠幩偟偨梒拵偺RNA偐傜cDNA傪崌惉偟丄娭楢堚揱巕偺敪尰傪掕検揑PCR朄偵傛傝夝愅偟偨丅斾妑幚尡偲偟偰丄僋儕僾僩價僆僔僗偺桿摫偐傜慼惗傑偱偺夁掱偵偮偄偰摨條偺挷嵏傪峴偭偨丅僈儞儅慄徠幩偵傛傞DNA偺僟儊乕僕偼丄摨慄検偺4He僀僆儞傪徠幩偟偨応崌偵斾傋偰寉旝偱偁傝丄24帪娫埲撪偵懳徠嬫(旕徠幩梒拵)偲摨儗儀儖偵傑偱夞暅偟偨丅4He僀僆儞偍傛傃僈儞儅慄傪徠幩偟偨僱儉儕儐僗儕僇梒拵偱偼丄偦偺僗僩儗僗偵墳偠偰DNA廋暅峺慺(Rad23, Rad51)偺敪尰傪桿摫偝傟偰偄偨丅堦曽丄4He僀僆儞偱偼妶惈巁慺傪彍嫀峺慺乮僇僞儔乕僛丄SOD摍乯偺敪尰偼曄壔偑柍偐偭偨偑丄僈儞儅慄徠幩屻俁乣12帪娫偵偐偗偰峈巁壔峺慺偺敪尰偑挊偟偔忋徃偟偨(恾俁)丅偙偺僈儞儅慄摿堎揑側墳摎偼丄峈巁壔峺慺偺敪尰桿摫偵妶惈巁慺偑夘嵼偟偰偄傞偙偲傪帵嵈偟偰偄傞丅堦曽丄DNA廋暅偵娭傢傞Rad23偺偍傛傃 Rad51偺敪尰偼僈儞儅慄偲4He僀僆儞曻幩慄偺椉曽偱桿摫偝傟偨偙偲偐傜峈巁壔峺慺偲偼堎側傞惂屼宯偑懚嵼偡傞偲峫偊傜傟偨丅僋儕僾僩價僆僔僗夁掱偱偺Rad51(憡摨慻姺偵傛傞廋暅峺慺)偺敪尰僷僞乕儞偼丄姡憞夁掱偍傛傃嵞悈榓捈屻偺嵺偵DNA擇廳嵔偺愗抐偑惗偠偰偄傞帠偑帵嵈偝傟偨丅幚嵺丄嵞悈榓捈屻偵妶惈巁慺偑尠挊偵敪惗偡傞帠偑帀幙偍傛傃 DNA偺巁壔偺柶塽慻怐妛揑夝愅偐傜徹柧偝傟偨丅姡憞偍傛傃曻幩慄僗僩儗僗乮70 Gy乯偵傛傞傾億僩乕僔僗偼娤嶡偝傟偢丄幚嵺傾億僩乕僔僗桿摫偵娭傢傞堚揱巕偺忋徃偼擣傔傜傟側偐偭偨丅姡憞偍傛傃曻幩慄僗僩儗僗偵敽偆DNA懝彎媦傃廋暅傗丄偦傟偵娭傢傞堚揱巕敪尰僷僞乕儞偺惉壥偼 PLoS ONE乮IF: 4.3)偵搳峞偟丄庴棟偝傟偨丅
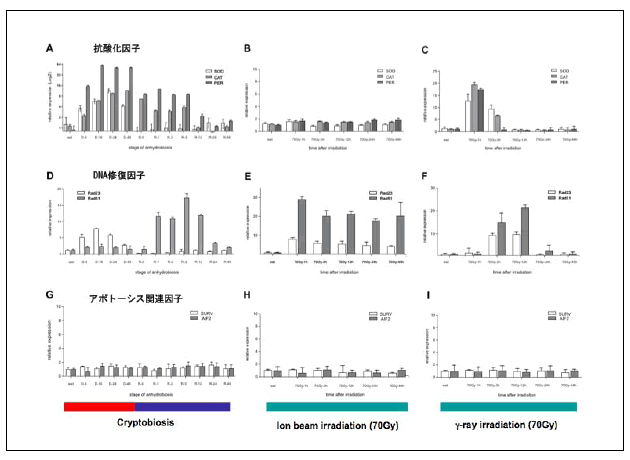
恾俁丂僱儉儕儐僗儕僇偺姡憞偲曻幩慄僗僩儗僗偵敽偆DNA偺懝彎偲廋暅偵娭傢傞堚揱巕偺敪尰僷僞乕儞丂A, D, G : 僋儕僾僩價僆僔僗嬫丄B, E, H : 4He僀僆儞價乕儉徠幩嬫丄C, F, I : 兞慄徠幩嬫
俁丏崱屻偺揥朷
丂僱儉儕儐僗儕僇悈榓梒拵偵兞-慄偍傛傃4He僀僆儞價乕儉傪偝傑偞傑側慄検偱徠幩偟丄RNA傪扨棧偟偰暯嬒壔cDNA儔僀僽儔儕乕偲捠忢偺cDNA儔僀僽儔儕乕傪怴偨偵峔抸偟偨丅偙偺摑崌EST僨乕僞儀乕僗偼僴僂僗僉乕僺儞僌堚揱巕偺傒側傜偢丄揮幨場巕傗僔僌僫儖揱払場巕側偳偺斾妑揑敪尰検偺彮側偄堚揱巕偑悢懡偔娷傑傟偰偄偨丅偙偺EST僨乕僞儀乕僗傪傕偲偵DNA僠僢僾傪嶌惉偟丄儅僀僋儘傾儗僀夝愅傪恑傔偰偄傞丅摿偵峈巁壔偵娭傢傞暘巕婡彉偺棟夝偑壛懍壔偝傟丄曻幩慄杊岇偵娭傢傞怴偨側庤朄偺峔抸偵戝偒側恑揥偑婜懸偝傟傞丅
係丏嶲峫暥專
Watanabe et al. (2006a) Int. J. Radiat. Biol. 82: 587.
Watanabe et al. (2006b) Int. J. Radiat. Biol. 82:835.
Nakahara et al., (2008) J. Insect Physiol. 54:1220.
Cornette et al., (2010) JBC 285:35889
Gusev et al., (2010) PLoS ONE (in press)